
Isaac Bianco and Stephen Wilson
Original paper reference:
Brain asymmetry is encoded at the level of axon terminal morphology
We use the left and right sides of our brains in different ways and this lateralisation of neural processing is observed throughout the animal kingdom. For instance, in humans, many aspects of the perception and generation of language occur predominantly in the left cerebral hemisphere and in many vertebrate species the left and right visual fields are specialised for processing different types of visual scenes. Presumably, these asymmetries in processing function are associated with left-right differences in the underlying neural architecture wherein the two sides of the brain are specialized for particular computational tasks. Although neuroanatomical asymmetries have been described at the level of differences in overall size, gross connectivity between brain regions and in the types of neurotransmitters used by neurons in equivalent regions on the left and right sides of the brain, very little is known about asymmetries in the microarchitecture of defined neuronal circuits that have distinct information processing roles. One reason for this is that such differences are likely to be relatively subtle. For instance, there could be differences in dendritic and/or axonal arbors - cellular processes by which neurons receive or transmit information to or from other neurons in the circuit, or in the synaptic connections that are formed on the arbors.
In this study, we resolved asymmetry at single-neuron resolution in a structure in the larval zebrafish brain called the habenulo-interpeduncular pathway and identified a previously unrecognized strategy by which neural circuits on the left and right sides of the brain can become distinct. We focused on this particular pathway because asymmetries between the left and right habenular nuclei are amongst the most conserved and conspicuous in the vertebrate brain. Furthermore, we had previously determined that the left and right habenulae establish laterotopic connections with their target, the interpeduncular nucleus (IPN), which lies at the midline of the ventral midbrain: whilst most left habenular axons innervate the dorsal part of the IPN, the majority of right habenular axons connect to the ventral IPN (Aizawa et al, 2005)
In collaboration with Jon Clarke (now at King's College London), we optimised a focal electroporation technique that allows nucleic acids or dyes to be introduced into individual cells in living zebrafish embryos. By expressing a membrane-tethered fluorescent protein, GFP, we were able to visualize the entire morphology of single habenular projection neurons in the intact brain (Fig 1A).
Figure 1 : Two subtypes of habenular projection neuron revealed by focal electroporation. (A) This panel shows an intact fish brain in which the entire morphology of a single right-sided habenular neuron is revealed following electroporation with a construct driving expression of GFP that localizes to the membrane of the neuron. (B and movie 1) An example of a L-typical axonal arbor, formed by most left habenular neurons and only a few right-sided cells. (C and movie 2) An example of a R-typical arbor elaborated by the majority of right habenular neurons. All panels show dorsal views, anterior top Scale bar in (A): 100µm.

Figure 1 : Two subtypes of habenular projection neuron revealed by focal electroporation. (A) This panel shows an intact fish brain in which the entire morphology of a single right-sided habenular neuron is revealed following electroporation with a construct driving expression of GFP that localizes to the membrane of the neuron. (B and movie 1) An example of a L-typical axonal arbor, formed by most left habenular neurons and only a few right-sided cells. (C and movie 2) An example of a R-typical arbor elaborated by the majority of right habenular neurons. All panels show dorsal views, anterior top Scale bar in (A): 100µm.
Movies 1- Movies 2
This led to the identification of two sub-types of habenular projection neuron which have axon terminal arbors with very distinct morphologies. The vast majority of left-sided habenular neurons elaborate axon arbors of one type, which we have called "L-typical" (Fig 1B). These arbors are shaped like a domed crown, localize to the dorsal part of the IPN and extend over a relatively large depth. Such arbors are also formed by a small number of right-sided cells. However, most right habenular neurons instead elaborate "R-typical arbors" which take the form of a flattened spiral and innervate the ventral IPN (Fig 1C). This axon arbor sub-type is also formed by a minority of left habenular cells.
Thus we have identified two neuronal subtypes that display distinct morphologies and connectivity and that are generated on both sides of the brain but in very different numbers. This represents a novel strategy for lateralisation of neuronal circuitry: the same (or very similar) circuit components are generated on both sides of the brain, but differences in the relative ratios of these shared components results in left-right specialization of circuit microarchitecture (Fig 2).
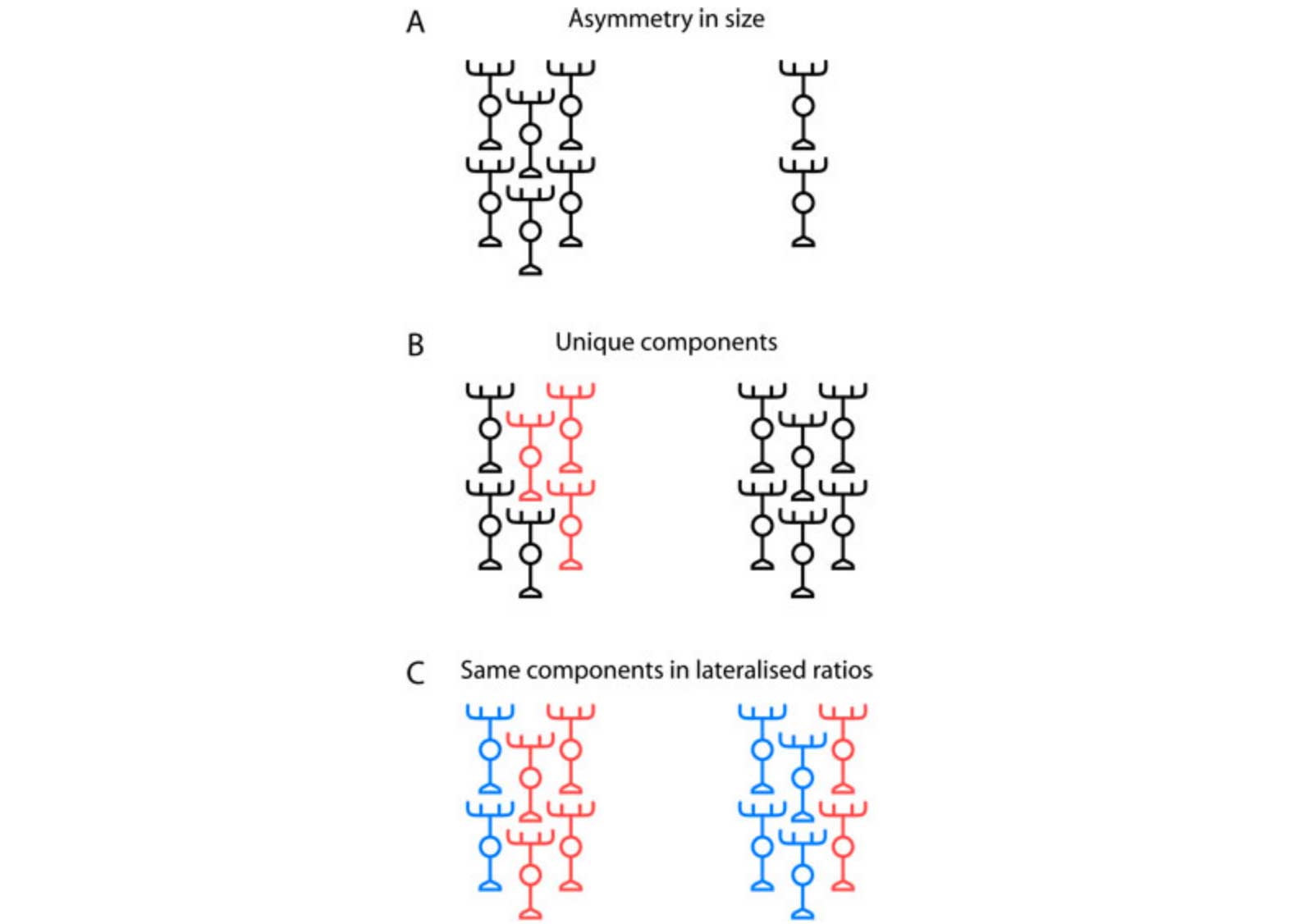
Figure 2: Models for lateralization of neural circuits. (A) In a simple 'scaling' model, equivalent regions on the left and right sides of the brain have the same composition and differ only in overall size. Cortical territories and language-associated fibre tracts appear to show this mode of lateralization. (B) Unique types of neuron or patterns of connectivity might be specified on one side of the brain (indicated by the unique red neurons on the left in this schematic). For example, the parapineal nucleus of neurons is located on the left side of the brain and innervates the left habenula with no equivalent neurons on the right. (C) The same circuitry components are produced on both sides of the brain but in different relative ratios so as to produce unique, lateralized circuits. This is the model we propose for habenulo-interpeduncular circuitry.
How do these different neuronal subtypes contribute to asymmetric processing in the habenulo-interpeduncular pathway? To try to shed light on this question we again turned to focal electroporation and examined the morphology of neurons in the target IPN. We found that whilst some post-synaptic neurons have their dendritic arbors completely confined to either the dorsal or ventral regions of the IPN neuropil (and as such will receive either L-typical or R-typical inputs), others types have dendritic arbors in both sub-regions. This suggests the IPN could integrate inputs from both sides of the brain whilst also containing circuits that preferentially relay left-dominant or right-dominant information to downstream targets.
To understand how asymmetric circuitry develops in the vertebrate brain, we examined the role of the parapineal, a small cluster of neurons that migrates towards and innervates the left habenula and is important for the expression of many of the molecular-genetic asymmetries between the left and right habenulae (see publications page for Concha 2003). In larvae in which the parapineal was removed by laser ablation, both habenulae predominantly innervated the ventral IPN, yielding a connectivity pattern that is superficially 'double right-sided'. However, examination of single neuron morphologies showed that subtle left-rght differences persist in parapineal-ablated embryos, suggesting that other, as yet unidentified developmental signals act to lateralise this highly conserved circuit.
Additional links
- This study has been shortlisted for the Biology Prize as part of the annual BioMed Central Research Awards
- Faculty of 1000
- UCL News
If you would like to read more about our asymmetry research, please visit our asymmetry research web pages. If you have any questions about this work, please contact Isaac (i.bianco@ucl.ac.uk) or Steve (s.wilson@ucl.ac.uk).
This project was funded by the Wellcome Trust and an European Communities grant entitled "Evolution and Development of Cognitive, Behavioural and Neural Lateralisation"